




Eme
| Eme | |
|---|---|
 | |
| Nome IUPAC | |
| 3-[18-(2-carboxyethyl)-8,13-bis(ethenyl)-3,7,12,17-tetramethylporphyrin-21,24-diid-2-yl]propanoic acid;iron(2+)[1] | |
| Caratteristiche generali | |
| Formula bruta o molecolare | C34H32FeN4O4[1] |
| Massa molecolare (u) | 616,5[1] |
| Numero CAS | |
| DrugBank | DBDB03014 |
| Indicazioni di sicurezza | |
L'eme (o gruppo eme) è un complesso chimico membro di una famiglia di composti chiamati porfirine contenente un atomo di ferro. L'eme costituisce il gruppo prostetico, cioè la parte non proteica, di una serie di proteine tra cui l'emoglobina, la mioglobina e i citocromi. Questa molecola deve la sua importanza al fatto che può legare l'ossigeno, sia in forma molecolare sia in altri composti (diossido di carbonio, monossido di carbonio CO, H2O, ecc.) proprio grazie all'atomo di ferro.
Struttura
[modifica | modifica wikitesto]L'eme è costituito da quattro anelli, interni all'emoglobina, pirrolici legati tra loro da ponti metinici (-CH=) formando un anello tetrapirrolico. A questa struttura sono legati quattro gruppi metilici (-CH3), due vinilici (CH2=CH-) e due propanoilici (-CH2-CH2-COO-). L'atomo di ferro viene legato alla fine ai quattro atomi di azoto disposti al centro dell'anello. Il ferro a sua volta può formare altri due legami, uno su ciascun lato dell'eme (chiamati quinta e sesta posizione di coordinazione). Nell'emoglobina e nella mioglobina la quinta posizione è occupata da una istidina.
L'eme può presentarsi sotto tre diverse forme: eme a, eme b ed eme c, dove l'eme b è il più comune dei tre; in particolare:
- eme a: differisce dall'eme b in quanto ha un gruppo metile ossidato a un gruppo formile (CHO), e una delle catene viniliche è rimpiazzata da una isoprenica; tra gli enzimi contenenti questo tipo di eme vi è la citocromo-c ossidasi;
- eme b: è presente nell'emoglobina e nella mioglobina;
- eme c: è simile all'eme b con la differenza che con le due catene viniliche è covalentemente legata alla sua apoproteina (parte proteica di una proteina[2]); tra le proteine che possiedono questa forma vi è il citocromo c.
- Forme dell'eme
-
 eme a
eme a -
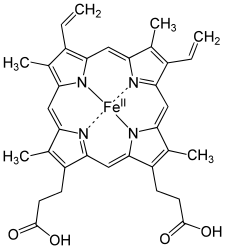 eme b
eme b -
 eme c
eme c



Funzione
[modifica | modifica wikitesto]La funzione di questa molecola è basata sull'atomo di ferro, in grado di legare in maniera reversibile l'ossigeno, utilizzandolo per trasportare elettroni nella catena respiratoria, nel ridurre specie reattive dell'ossigeno (catalasi e perossidasi) o semplicemente trasportandolo nel sangue (emoglobina) o immagazzinandolo nei muscoli (mioglobina). Poiché nelle cellule il principale sito di ossidazione sono i mitocondri, molti enzimi contenenti eme si trovano lì.
Il ferro può esistere negli stati di ossidazione +2 e +3, ma solo lo stato di ossidazione minore è in grado di legare l'ossigeno. Quando il ferro si trova nello stato di ossidazione +3 non è più in grado di legare l'ossigeno e l'emoglobina prende il nome di metaemoglobina. La capacità di legare l'ossigeno da parte dell'eme così come quella di trattenere gli elettroni vengono modificate dalla proteina nella quale si trova.
Sintesi
[modifica | modifica wikitesto]
La sintesi dell'eme ha luogo nel citoplasma e nel mitocondrio della cellula. Il processo enzimatico che produce l'eme va sotto il nome di sintesi porfirinica, poiché l'eme fa parte della famiglia delle porfirine. Questo processo di sintesi, e gli enzimi che ne fanno parte, sono molto conservati tra le varie specie. Nell'uomo, questa via di sintesi serve quasi esclusivamente a produrre eme; in altre specie serve anche a produrre sostanze simili come la cobalamina (vitamina B12). La sintesi dell'eme ha inizio dalla sintesi di acido (delta)-aminolevulinico dall'amminoacido glicina e dal Succinil CoA proveniente dal ciclo di Krebs (ciclo dell'acido citrico). L'attività dell'enzima responsabile di questa reazione (ALA sintasi) è regolato dal livello intracellulare di ferro e dalla concentrazione di eme. Gli organi principalmente coinvolti nella sintesi dell'eme sono il fegato e il midollo osseo, sebbene tutte le cellule lo richiedano per funzionare bene. Gli eritrociti a differenza dei loro precursori, i reticolociti, non possono produrlo.
Nel processo interviene l'enzima uroporfirinogeno III sintasi.
Degradazione
[modifica | modifica wikitesto]L'eme libero ha il principale effetto di indurre la formazione di ROS (specie reattive dell'ossigeno), provocando danno ossidativo all'ambiente. Pertanto è necessario che la cellula catabolizzi rapidamente le specie di scarto.
Dopo l'emolisi, l'emoglobina liberamente rilasciata nel sangue viene dapprima captata dall'aptoglobina (Hp), formando il complesso Hb/Hp, che viene endocitato dalle cellule del sistema reticoloendoteliale via il trasportatore CD163 (la componente proteica viene degradata ad amminoacidi e il gruppo prostetico viene liberato). Quando la disponibilità di Hp va incontro a esaurimento, l'eme viene rilasciato dall'impalcatura proteica, e quindi captato dall'albumina, da HDL/LDL o dall'emopessina (Hx), che analogamente a Hp contribuisce ad accrescere il pool di eme libero nelle cellule adibite al suo catabolismo, attraversando il trasportatore CD91.[3]
L'eme libero viene degradato dall'enzima eme ossigenasi (HO-1/2) che porta alla formazione di bilirubina, un potente agente antiossidante di colore giallognolo. L'eme dei globuli rossi viene invece degradato nella milza. Più correttamente, dall'apertura dell'anello porfirinico si origina prima biliverdina e ossido di carbonio, con rilascio dell'atomo di ferro centrale; la biliverdina, poi, viene convertita a bilirubina dall'enzima biliverdina reduttasi.
Notare che dalla degradazione dell'anello viene generato ossido di carbonio, noto gas tossico che ha elevata affinità per l'eme stesso. La reale funzione di questo gas nel sito di produzione è, invece, modulare delle risposte metaboliche postume al catabolismo dell'eme, come la respirazione cellulare (citocromo mitocondriale) e la produzione del secondo messaggero GMP ciclico.
La bilirubina presente nel sangue viene captata dal fegato, coniugata con acido glucuronico e quindi espulsa con la bile nel tratto intestinale. Qui si trasforma nei bilinogeni incolori (urobilinogeno, mesobilirubinogeno e stercobilinogeno) e quindi in biline (urobilina e stercobilina), responsabili della colorazione delle feci.
Malattie metaboliche
[modifica | modifica wikitesto]Difetti degli enzimi responsabili della sintesi o della degradazione dell'eme sono responsabili di disordini ereditari o acquisiti, chiamati porfirie.
Note
[modifica | modifica wikitesto]- ^ a b c (EN) PubChem, Protoheme, su pubchem.ncbi.nlm.nih.gov, National Center for Biotechnology Information. URL consultato il 23 novembre 2021.
- ^ apolipoproteina (o apoproteina) in "Dizionario di Medicina", su treccani.it. URL consultato il 23 novembre 2021.
- ^ Larsen R., Gouveia Z., Soares M., Gozzelino R. (2012). Heme cytotoxicity and thepathogenesis of immune-mediated inflammatory diseases. Frontiers In Pharmacology.
Bibliografia
[modifica | modifica wikitesto]- Stryer L., Biochimica, quarta edizione (codice ISBN 88-08-09806-0)
Voci correlate
[modifica | modifica wikitesto]Collegamenti esterni
[modifica | modifica wikitesto]- eme, su Treccani.it – Enciclopedie on line, Istituto dell'Enciclopedia Italiana.
- (EN) heme, su Enciclopedia Britannica, Encyclopædia Britannica, Inc.
| Controllo di autorità | Thesaurus BNCF 42237 · LCCN (EN) sh85060147 · GND (DE) 4244498-6 · BNF (FR) cb119813059 (data) · J9U (EN, HE) 987007555531005171 |
|---|