





Dimetrodon
| Dimetrodon | |
|---|---|
 | |
| Intervallo geologico | |
| Stato di conservazione | |
Fossile | |
| Classificazione scientifica | |
| Dominio | Eukaryota |
| Regno | Animalia |
| Phylum | Chordata |
| Superclasse | Tetrapoda |
| Classe | Synapsida |
| Ordine | † Pelycosauria |
| Famiglia | † Sphenacodontidae |
| Sottofamiglia | † Sphenacodontinae |
| Genere | † Dimetrodon Cope, 1878 |
| Nomenclatura binomiale | |
| † Dimetrodon limbatus Cope, 1878 (originariamente Clepsydrops limbatus) | |
| Sinonimi | |
| |
| Specie | |

Dimetrodon (il cui nome significa "denti di due misure"), è un genere estinto di sinapside sfenacodontide vissuto nel periodo Permiano, età del Cisuraliano, circa 295-272 milioni di anni fa (Sakmariano-Kunguriano).[1][2][3] Si tratta di un membro della famiglia degli Sphenacodontidae. La caratteristica più famosa di Dimetrodon è senza dubbio la grande vela dorsale, formata da processi spinali allungati verticalmente collegati tra loro tramite uno strato di pelle. L'animale era quadrupede e aveva un cranio alto e curvo con grandi denti di diverse dimensioni posti lungo le mascelle. La maggior parte dei fossili è stata ritrovata nel sud-ovest degli Stati Uniti, e la maggior parte di essi proviene da un deposito geologico chiamato Red Beds, del Texas e dell'Oklahoma. Più recentemente, fossili di Dimetrodon sono stati ritrovati anche in Germania. Fin dalla sua descrizione nel 1878, sono state descritte oltre una dozzina di specie ascritte al genere.
Sebbene nella cultura popolare venga spesso scambiato per un dinosauro o come loro contemporaneo, Dimetrodon si estinse circa 40 milioni di anni prima dell'apparizione dei dinosauri. Sebbene il suo aspetto e la sua fisiologia lo facciano assomigliare ai rettili, il Dimetrodon è più strettamente legato ai mammiferi, anche se non è un diretto antenato.[2] Il Dimetrodon è stato catalogato come un "proto-mammifero" (stem-mammal).[2] Insieme ai mammiferi questo gruppo, che contiene Dimetrodon, compone il clado (gruppo evolutivo) chiamato Synapsida, mentre dinosauri, rettili e uccelli fanno parte del clado separato, Sauropsida. Le singole aperture nel cranio dietro ogni occhio, conosciute come finestre temporali, e altre caratteristiche del cranio distinguono Dimetrodon e i mammiferi dalla maggior parte dei primi sauropsida.
Dimetrodon era, probabilmente, uno dei superpredatori degli ecosistemi del Cisuraliano, cacciando e nutrendosi di pesci e tetrapodi, tra cui rettili e anfibi, mentre le specie più piccole di Dimetrodon potrebbero aver avuto ruoli ecologici diversi. Sono state avanzate numerose ipotesi sulla funzione della vela di Dimetrodon: l'ipotesi più accreditata fin dalla sua scoperta è che questa struttura servisse per la termoregolazione corporea e servisse ad assumere e disperdere calore più velocemente. Tuttavia, la scoperta dell'endotermia negli sfenacodontidi[4] ha smontato tale ipotesi, e ora si pensa principalmente che la vela avesse solo la funzione di richiamo sessuale, così come in altri animali come Sphenacodon e Edaphosaurus.
Descrizione
[modifica | modifica wikitesto]
Dimetrodon era un sinapside quadrupede, le cui varie specie variavano per dimensioni, con una lunghezza dai 1,7 metro a 4,6 metri (5,6 a 15,1 piedi) e si stima che il suo peso si aggirasse tra i 28 e i 250 chilogrammi (62 e 551 libbre).[5] La più grande specie conosciuta di Dimetrodon è D. angelensis di 4,6 metri (15 piedi), mentre la specie più piccola è D. teutonis di soli 60 centimetri (24 pollici).[6][7] Le specie più grandi di Dimetrodon dovevano essere i più grandi predatori del Permiano inferiore, anche se l'affine Tappenosaurus, conosciuto solo per frammenti scheletrici ritrovati in rocce un po' più recenti, potrebbe essere stato ancora più grande con una lunghezza di circa 5,5 metri (18 piedi).[8][9] Tuttavia alcune specie di Dimetrodon sono conosciute solo da esemplari giovani quindi gli animali adulti potrebbero essere stati ben più grandi.[10]
Cranio
[modifica | modifica wikitesto]Dimetrodon possedeva un'unica grande apertura su entrambi i lati della parte posteriore del cranio, il che lo collega con i mammiferi e lo distingue dalla maggior parte dei primi sauropsidi, che mancavano completamente di aperture o ne avevano due. Caratteristiche come le creste sulla parte interna della cavità nasale e una cresta sul retro della mandibola sono pensate per essere parte di una progressione evolutiva dei primi tetrapodi (vertebrati a quattro zampe) verso i mammiferi.
Il cranio di Dimetrodon è alto, curvo e compresso lateralmente. Le orbite sono posizionate in alto e dietro ogni cavità oculare vi è un unico foro, chiamato finestra temporale. Un foro aggiuntivo nel cranio, la finestra sovratemporale, può essere vista dall'alto. La parte posteriore del cranio (la regione occipitale) è orientata con una leggera angolazione verso l'alto, una caratteristica che condivide con tutti gli altri primi sinapsidi.[11] Il margine superiore del cranio declivi in un arco convesso alla punta del muso. La punta della mascella superiore, formata dalla premascella, è sollevata sopra la parte della ganascia formata dall'osso mascellare, venendo divisa da una sorta di solco che divide la premascella dalla mascella vera e propria. Questo solco viene chiamato diastema, o lacuna nella fila dei denti. Il cranio di Dimetrodon è inoltre più robusto di quello di un dinosauro, proprio grazie all'assenza di cavità.
-
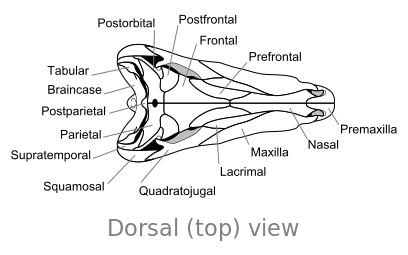 Ossa del cranio di Dimetrodon, in vista dorsale
Ossa del cranio di Dimetrodon, in vista dorsale -
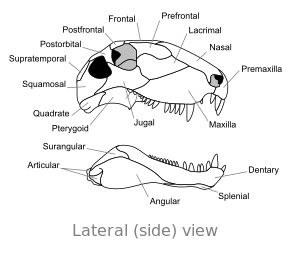 Ossa del cranio di Dimetron, in vista laterale
Ossa del cranio di Dimetron, in vista laterale -
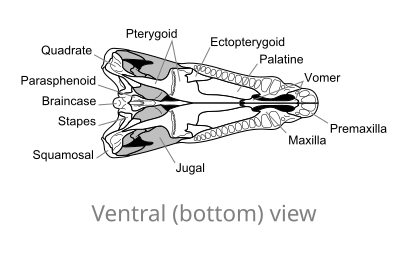 Ossa del cranio di Dimetrodon, in vista ventrale
Ossa del cranio di Dimetrodon, in vista ventrale -
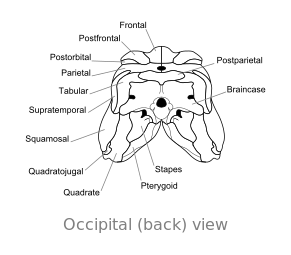 Ossa del cranio di Dimetrodon, in vista occipitale
Ossa del cranio di Dimetrodon, in vista occipitale




Denti
[modifica | modifica wikitesto]
Le dimensioni dei denti variano in base alla lunghezza delle ganasce, dando così a Dimetrodon il suo nome, che significa infatti "denti di due misure" in riferimento ai gruppi di piccoli e grandi denti.[12] Una o due coppie di denti caniniformi (denti dalla grande punta simili ai canini) si estendono dalla mascella. Sono presenti anche grandi incisivi caniniformi presenti all'estremità della mascella superiore e inferiore, radicati nella premascella e anche nell'osso dentario. I denti più piccoli erano presenti lungo tutta la mascella e dietro i denti caniniformi, diventando più piccolo man mano che si retrocedeva nella mascella.[13]
Molti denti sono più ampi nella loro sezione e si restringono vicino alle ganasce, dando loro l'aspetto di una lacrima. La forma "a lacrima" dei denti sono unici di Dimetrodon e altri sfenacodontidi strettamente correlati, e spesso aiutano i paleontologi a distinguerli da altri primitivi sinapsidi.[7] Come in molti altri sinapsidi primitivi, i denti della maggior parte delle specie di Dimetrodon sono dentellati sui bordi.[7] Le dentellature dei denti di Dimetrodon erano così fini che sembravano piccole crepe.[14] Anche il dinosauro Albertosaurus aveva dentellature simili, ma, alla base di ogni dentellatura vi è un tondo vuoto, che sarebbe servito per distribuire la forza su una grande area superficiale e prevenire le sollecitazioni e diffonderle attraverso il dente. A differenza di Albertosaurus, però, i denti di Dimetrodon mancavano di adattamenti che fermavano la formazione di crepe nelle loro scanalature.[14] I denti della specie D. teutonis mancavano di dentellature, ma avevano ancora i bordi taglienti.[14]
Uno studio del 2014 mostra che i denti del genere Dimetrodon erano adattati alle dimensioni delle sue prede. Le specie più piccole, come D.milleri non avevano dentellature in quanto mangiava piccole prede. Più la preda era grande, più i denti delle varie specie di Dimetrodon sviluppavano dentellature e un progressivo aumento in termini di dimensioni. La specie D. limbatus aveva dentellature di smalto che aiutavano a sminuzzare la carne. Le stesse dentellature si possono trovare anche in Secodontosaurus. La seconda specie più grande di Dimetrodon, D. grandis, aveva dentellature simili a quelle degli squali e dei dinosauri teropodi, rendendo i suoi denti ancora più specializzati per tagliare la carne. Questo studio non solo mostra l'evoluzione di Dimetrodon nel corso di milioni di anni, ma dimostra anche che Dimetrodon era in continua corsa agli armamenti contro le sue prede. Come la preda diveniva più grande, più il Dimetrodon contrastata crescendo in dimensioni e sviluppando denti più taglienti.[15]
Cavità nasale
[modifica | modifica wikitesto]Sulla superficie interna della sezione nasale del cranio erano presenti piccole strutture chiamate creste nasoturbinali, probabilmente adibite a sostenere delle strutture cartilagine, che aumentato la superficie dell'epitelio olfattivo, lo strato di tessuto che rileva gli odori. Queste creste sono molto più piccole di quelle dei sinapsidi più evoluti, alla fine del Permiano e del Triassico, i cui nasoturbinali erano una prova di endotermia, perché sostenevano le mucose che scaldavano e umidificavano l'aria in entrata. Così, la cavità nasale di Dimetrodon sembra essere una transizione tra quella dei primi vertebrati terrestri e i mammiferi.[16]
Giunto mandibolare e orecchio
[modifica | modifica wikitesto]Un'altra caratteristica di transizione di Dimetrodon è una cresta nella parte posteriore della mandibola chiamato lamina riflessa. La lamina riflessa viene trovata sulle articolazioni delle osso, che si collega alla osso quadrato del cranio per formare la mandibola. Gli antenati dei mammiferi successivi, hanno un'articolazione e un osso quadrato separato dalla mandibola mentre l'articolazione si sviluppa nel martello dell'orecchio medio. La lamina riflessa fa parte di un anello chiamato anulus timpanico che supporta il timpano in tutti i mammiferi viventi.[17]
Coda
[modifica | modifica wikitesto]
La coda di Dimetrodon costituisce gran parte della lunghezza totale del corpo e comprende circa 50 vertebre caudali. I primi scheletri di Dimetrodon era incompleti e la coda mancava; l'unico indizio della sua presenza era la scoperta di undici vertebre caudali ritrovate vicino ad un fianco. Dal momento che queste prime vertebre caudali si restringevano rapidamente man mano che si allontanavano dal fianco, molti paleontologi alla fine del XIX secolo e all'inizio del XX pensavano che Dimetrodon avesse una coda molto corta. Solo dopo il 1927, fu finalmente ritrovata e descritta una coda in gran parte completa di Dimetrodon.[18]
Vela
[modifica | modifica wikitesto].jpg)
La vela di Dimetrodon è formata da un allungamento delle spine neurali sporgenti dalle vertebre. Ogni spina neurale varia in sezione e forma dalla sua base alla sua punta in ciò che è noto come differenziazione "dimetrodonte".[19] Vicino al corpo della vertebra, la sezione trasversale della colonna vertebrale è compressa lateralmente in una forma rettangolare, e più vicino alla punta, esso assume una forma ad otto come una scanalatura che corre lungo entrambi i lati della colonna vertebrale. Si pensa che la forma ad otto servisse per rafforzare la colonna vertebrale, impedendo flessione e fratture.[20] Una sezione trasversale della colonna vertebrale di un esemplare di Dimetrodon giganhomogenes è di forma rettangolare, ma conserva la forma di otto anelli sagomati vicino al suo centro, indicando che la forma delle spine poteva cambiare da individuo ad individuo o in base all'età.[21] L'anatomia microscopica di ogni colonna vertebrale varia dalla base alla punta, il che indica dove si ancoravano i muscoli della schiena e dove era esposta parte della vela. La porzione più bassa, o prossimale, della colonna vertebrale presenta una superficie ruvida che sarebbe servita come punto di ancoraggio per i muscoli epaxiali della schiena, e ha anche una rete di tessuti connettivi chiamati fibre di Sharpey che indicano, che era incorporata al corpo. Più in alto sulla parte distale (esterna) della colonna vertebrale, la superficie ossea è liscia. Il periostio, uno strato di tessuto che circonda l'osso, è coperto di piccole scanalature che presumibilmente supportavano i vasi sanguigni che vascolarizzavano la vela.[22]
I grandi solchi che correvano lungo tutta la lunghezza della colonna vertebrale, una volta si pensava fossero un canale per i vasi sanguigni, ma dal momento che l'osso non contiene canali vascolari, la vela non doveva essere così vascolarizzata come si pensava. Alcuni esemplari di Dimetrodon conservano aree deformate delle spine neurali che sembrano essere guarite dopo la frattura. L'osso corticale che cresce in queste aree deformate è altamente vascolarizzato, suggerendo che dei tessuti molli dovevano essere presente sulla vela per rifornire il sito di vasi sanguigni.[20] L'osso lamellare costituisce la maggior parte della sezione trasversale della colonna vertebrale neurale, e contiene le linee di arresto della crescita che possono essere utilizzate per determinare l'età di ogni individuo alla morte.[23] In molti esemplari di D. gigashomogenes le porzioni distali delle spine sono piegate bruscamente, indicando che la vela aveva un profilo irregolare quando l'animale era in vita. La loro flessibilità potrebbe suggerire che il tessuto molle che connetteva le spine neurali poteva non essere presente fino alle punte delle spine, il che significa che l'estensione della vela di pelle non era poi così ampia come si pensava.[19]
Pelle
[modifica | modifica wikitesto]Non è mai stata ritrovata alcuna prova fossile sulla pelle di Dimetrodon. Tuttavia è stata ritrovata un'impressione di pelle di un animale correlato a Dimetrodon,Estemmenosuchus, che possedeva una pelle rugosa e secca e fornita di ghiandole, simile a quella degli elefanti[24] con piccole strutture simili a squame sul ventre e sulla parte inferiore della coda, come gli altri sinapsidi.[25][26]
Scoperta e classificazione
[modifica | modifica wikitesto]Le prime descrizioni di Cope
[modifica | modifica wikitesto]
I fossili oggi attribuiti a Dimetrodon, furono studiati dal paleontologo americano Edward Drinker Cope, nel 1870. Cope aveva ottenuto i fossili insieme a quelli di molti altri tetrapodi del Permiano (vertebrati a quattro zampe), inviategli da diversi collezionisti che stavano esplorando un gruppo di rocce in Texas, chiamate Red Bed. Tra questi collezionisti vi erano il naturalista svizzero Jacob Boll, il geologo del Texas W.F. Cummins e il paleontologo amatoriale Charles Hazelius Sternberg.[27] La maggior parte dei campioni di Cope furono mandati all'American Museum of Natural History e/o al Museo Walker dell'Università di Chicago (la maggior parte della collezione dei fossili di Walker è ora ospitata al Field Museum).
Sternberg inviò alcuni dei suoi campioni al paleontologo tedesco Ferdinand Broili, presso l'Università di Monaco di Baviera, sebbene Broili non fosse così eccezionale quando si trattava di descrivere i campioni. Anche il rivale di Cope, Othniel Charles Marsh ritrovò alcune ossa di Dimetrodon, che inviò poi al Museo Walker.[28] Il nome Dimetrodon venne usato per la prima volta nella descrizione di Cope (1878), quando Cope denominò le prime tre specie, ossia Dimetrodon incisivus, D. rectiformis e D. gigas, sulla rivista scientifica Proceedings of the American Philosophical Society.[29]
Tuttavia, la prima descrizione dei fossili di Dimetrodon avvenne un anno prima, quando Cope diede il nome alla specie Clepsydrops limbatus, dai Red Bed del Texas.[30] In seguito, la specie venne cambiata da C. limbatus ad una specie di Dimetrodon, divenendo D. limbatus.
Con la sua descrizione di Clepsydrops natalis, Cope fu il primo a descrivere un sinapside crestato, sebbene descrisse la vela di Dimetrodon come una pinna e la confrontò con la cresta dorsale del moderno basilisco (gen. Basiliscus). Le vele non erano conservate negli esemplari di D. incisivus e D. gigas, come descritto da Cope (1878), ma lunghe spine allungate erano presenti nei fossili di D. rectiformis.[13] Cope commentò anche lo scopo della vela nel 1886, affermando che, "l'utilità [della vela] è difficile da immaginare. A meno che l'animale non abbia avuto abitudini acquatiche e nuotava sul dorso, la cresta o la pinna non poteva avere uno scopo motorio ... gli arti non erano abbastanza lunghi, e gli artigli non erano abbastanza affilati per dimostrare delle abitudini arboree, come nel genere esistente Basiliscus, dove è effettivamente presente una cresta simile ".[13]
Le prime descrizioni del XX secolo
[modifica | modifica wikitesto]Nei primi decenni del XX secolo, il paleontologo americano Ermine Cowles Case, autore di molti studi su Dimetrodon descrisse diverse nuove specie del genere. Case ricevette diversi finanziamenti dal Carnegie Institution per il suo studio si numerosi esemplari di Dimetrodon nelle collezioni del Museo Americano di Storia Naturale e in molti altri musei.[28] Molti di questi fossili erano stati raccolti da Cope, ma non erano ancora stati accuratamente descritti, in quanto Cope era noto per erigere nuove specie anche sulla base di pochi frammenti di ossa.
A partire dalla fine del 1920, il paleontologo Alfred Romer ristudiò molti esemplari di Dimetrodon e diede il nome a diverse nuove specie. Nel 1940, Romer fu coautore dell'ampio studio con Llewellyn Ivor Price, definito "Revisione di Pelycosauria", in cui le specie di Dimetrodon sono state rivalutate da Case. La maggior parte dei nomi delle specie considerate valide da Romer e Price sono ancora oggi in uso.[22]
Nuovi esemplari
[modifica | modifica wikitesto]Nei decenni seguenti: alla monografia di Romer e Price vennero aggiunte numerose nuove specie di Dimetrodon, provenienti da località anche al di fuori del Texas e dell'Oklahoma. Il primo fu ritrovato nel Four Corners, nello Utah, nel 1966[31] mentre un altro fu ritrovato in Arizona, nel 1969.[32] Nel 1975, Olson riferì del nuovo materiale rinvenuto nell'Ohio, a Dimetrodon.[33] Una nuova specie di Dimetrodon, chiamata D. occidentalis (il cui nome significa "dimetrodonte occidentale") è stata denominata nel 1977, dal Nuovo Messico.[34] Gli esemplari ritrovati nello Utah e in Arizona, probabilmente appartengono alla specie D. occidentalis.[35]
Prima di queste scoperte, esisteva una teoria secondo cui nel Permiano inferiore, un mare interno separava quello che oggi sono il Texas e l'Oklahoma dalle terre più occidentali, isolando Dimetrodon in una piccola regione del Nord America, mentre lo sphenacodontide più piccolo Sphenacodon, dominava la zona occidentale. Sebbene la presenza di questo mare interno sia molto probabile, la scoperta di fossili al di fuori del Texas e dell'Oklahoma mostrano che la sua estensione era limitata e che non era una barriera efficace per la distribuzione di Dimetrodon.[34][36]
Nel 2001, una nuova specie di Dimetrodon, chiamata D. teutonis, è stata descritta da una località risalente al Permiano inferiore, presso la foresta della Turingia, in Germania, ampliando la gamma geografica di Dimetrodon anche fuori dal Nord America per la prima volta.[5]
Specie
[modifica | modifica wikitesto]| Specie | Autore/i | Locazione | Stato | Sinonimi | Immagine |
|---|---|---|---|---|---|
| Dimetrodon angelensis | Olson, 1962 | Texas | Valida | 
| |
| Dimetrodon borealis | Leidy, 1854 | Isola del Principe Edoardo | Valida | Originariamente conosciuta come Bathygnathus borealis | 
|
| Dimetrodon booneorum | Romer, 1937 | Texas | Valida | ||
| Dimetrodon dollovianus | Case, 1907 | Texas | Valida | Embolophorus dollovianus Cope, 1888 | |
| Dimetrodon gigashomogenes | Case, 1907 | Texas | Valida | 
| |
| Dimetrodon grandis | Romer & Price, 1940 | Oklahoma Texas |
Valida | Clepsydrops gigas Cope, 1878 Dimetrodon gigas Cope, 1878 Theropleura grandis Case, 1907 Bathyglyptus theodori Case, 1911 Dimetrodon maximus Romer 1936 |

|
| Dimetrodon kempae | Romer, 1937 | Texas | Possibile nomen dubium | ||
| Dimetrodon limbatus | Romer & Price, 1940 | Oklahoma Texas |
Valida | Clepsydrops limbatus Cope, 1877 Dimetrodon incisivus Cope, 1878 Dimetrodon rectiformis Cope, 1878 Dimetrodon semiradicatus Cope, 1881 |
|
| Dimetrodon loomisi | Romer, 1937 | Texas Oklahoma |
Valida | 
| |
| Dimetrodon macrospondylus | Case, 1907 | Texas | Valida | Clepsydrops macrospondylus Cope, 1884 Dimetrodon platycentrus Case, 1907 |
|
| Dimetrodon milleri | Romer, 1937 | Texas | Valida | 
| |
| Dimetrodon natalis | Romer, 1936 | Texas | Valida | Clepsydrops natalis Cope, 1878 | 
|
| Dimetrodon occidentalis | Berman, 1977 | Arizona Nuovo Messico Utah |
Valida | ||
| Dimetrodon teutonis | Berman et al., 2001 | Germania | Valida |
Filogenesi
[modifica | modifica wikitesto]Dimetrodon è uno dei primi membri di un gruppo chiamato sinapsidi, che comprendono i mammiferi e molti dei loro parenti estinti, sebbene Dimetrodon non sia l'antenato di tutti i mammiferi (che sono apparsi milioni di anni più tardi[37]). Dimetrodon è spesso scambiato per un dinosauro nella cultura popolare, pur essendosi estinto circa 40 milioni di anni prima dell'apparizione dei primi dinosauri nel Triassico. Come tutti sinapsidi, Dimetrodon è più strettamente legato ai mammiferi che ai dinosauri o di qualsiasi altro rettile vivente. Dal 1900, la maggior parte dei paleontologi si riferisce a Dimetrodon come un rettile, in accordo con la tassonomia di Linneo, che ha classificato Reptilia come classe e Dimetrodon come un genere all'interno di questa classe. I mammiferi sono stati assegnati a una classe separata, e Dimetrodon è stato descritto come un "rettile simile ad un mammifero". I paleontologi hanno teorizzato che i mammiferi si siano evoluti da questo gruppo, in una transizione da rettile a mammifero.
Tassonomia dei sinapsidi
[modifica | modifica wikitesto]
Sotto la sistematica filogenetica, i discendenti dell'ultimo antenato comune di Dimetrodon e di tutti i rettili viventi, includono tutti i mammiferi, perché Dimetrodon è più strettamente imparentato ai mammiferi di qualsiasi altro rettile vivente. Così, se si desidera evitare il clado che contiene sia mammiferi e i rettili viventi, Dimetrodon non deve essere incluso in tale clado, né in alcun altro clado di "rettili simile ai mammiferi". I discendenti dell'ultimo antenato comune dei mammiferi e dei rettili (che è apparso circa 310 milioni di anni fa, nel Carbonifero superiore) sono quindi divisi in due cladi: Synapsida, che comprende Dimetrodon e i mammiferi, e i Sauropsida, che comprendono i rettili viventi e tutti i rettili estinti più strettamente correlati tra loro che con i mammiferi.[2]
All'interno del clado Synapsida, Dimetrodon fa parte del clado degli Sphenacodontia, che è stato proposto come un gruppo di sinapsidi, all'inizio del 1940 dai paleontologi Alfred Romer e Llewellyn Ivor Price, insieme al gruppo degli Ophiacodontia e degli Edaphosauria.[38] Tutti e tre i gruppi sono noti dal Carbonifero superiore al Permiano inferiore. Romer e Price li distinguono principalmente per le caratteristiche postcraniali, come le forme delle arti e delle vertebre. Gli Ophiacodontia sono considerati il gruppo più primitivo, perché i suoi membri erano più imparentati con i rettili, mentre gli Sphenacodontia erano il gruppo più avanzato poiché i suoi membri erano più imparentati con i Therapsida, che comprendeva i parenti più stretti dei mammiferi. Romer e Price hanno posizionati un altro gruppo tra i sinapsidi primitivi, chiamati varanopidi, all'interno di Sphenacodontia, considerandolo più primitivo rispetto ad altri sfenacodonti, come Dimetrodon.[39] In passato si pensava che i varanopidi fossero strettamente imparentati con gli sfenacodonti simili a Dimetrodon, in quanto entrambi i gruppi erano carnivori, anche se i varanopidi sono molto più piccoli e non possiedono più vele.
La visione moderna dei rapporti tra sinapsidi è stata proposta dal paleontologo Robert R. Reisz, nel 1986, il cui studio includono le caratteristiche per lo più trovate nel cranio, piuttosto che nello scheletro postcraniale.[40] Dimetrodon è ancora considerato uno sfenacodonte sotto questo filogenesi, ma ora i varanopidi sono considerati dei sinapsidi più basali, non conformi al clado degli Sphenacodontia. All'interno di Sphenacodontia vi è il gruppo Sphenacodontoidea, che a sua volta contiene Sphenacodontidae e Therapsida. Gli Sphenacodontidae è il gruppo che contiene Dimetrodon e diverse altri sinapsidi con la vela, come Sphenacodon, Ctenospondylus e Secodontosaurus, mentre Therapsida comprende i mammiferi e i loro parenti del Permiano e del Triassico.
Di seguito è riportato il cladogramma del clado di Synapsida, che segue la filogenesi di Synapsida modificata dall'analisi di Benson (2012):[39]
| Amniota |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Il cladogramma seguente mostra i rapporti di alcune specie di Dimetrodon, da Brink et al., (2015):[41]
| Sphenacodontidae |
| ||||||||||||||||||||||||||||||||||||
Paleobiologia
[modifica | modifica wikitesto]Funzione della vela
[modifica | modifica wikitesto]I paleontologi hanno proposto diverse ipotesi sull'effettiva funzione della vela in vita. Una delle prime ipotesi proposte fu il loro impiego nella mimetizzazione dell'animale, che poteva così mimetizzarsi tra i canneti, mentre Dimetrodon aspettava la preda. Un'altra ipotesi piuttosto fantasiosa è che essa funzionasse da vera e propria vela, per il movimento in acqua dell'animale, catturando il vento.[42] Un'altra ipotesi è che le lunghe spine neurali avessero la funzione di stabilizzatori del tronco limitando il movimento verticale, che avrebbe consentito un movimento laterale più efficace.[20]
Termoregolazione
[modifica | modifica wikitesto]Nel 1940, Alfred Romer e Llewellyn Ivor Price proposero che la vela avesse una funzione di termoregolazione, consentendo ai Dimetrodon di scaldare i loro corpi più efficacemente esponendoli al calore del sole. Negli anni successivi, sono stati creati numerosi modelli per valutare l'efficacia della termoregolazione dei Dimetrodon. Ad esempio, in un articolo del 1973, nella rivista Nature, i paleontologi CD Bramwell e PB Fellgett stimarono che un individuo di 200 kg (440 libbre) sarebbe dovuto rimanere per circa un'ora e mezza al sole per far alzare la sua temperatura corporea da 26 a 32 °C (79 a 90 °F).[43] Nel 1986, Steven C. Haack ha concluso che il riscaldamento dell'animale doveva essere ben più lento di quanto si pensasse e che per l'intero processo ci sarebbero volute, probabilmente, almeno quattro ore. Utilizzando un modello basato su una varietà di fattori ambientali e ipotizzate alcuni aspetti fisiologici di Dimetrodon, Haack ha scoperto che la vela avrebbe permesso a Dimetrodon di scaldarsi più velocemente la mattina e raggiungere una temperatura corporea leggermente più alto durante il giorno, ma la vela era inefficace nel rilasciare il calore in eccesso e non permetteva a Dimetrodon di mantenere una temperatura corporea più alta durante la notte.[44] Nel 1999, un gruppo di ingegneri meccanici ha creato un modello al computer per analizzare le capacità della vela nella regolazione della temperatura corporea durante le diverse stagioni, e ha concluso che la vela era uno strumento benefico sia per l'assorbimento sia per il rilascio di calore, in ogni momento durante l'anno.[45]

La maggior parte di questi studi danno due ruoli di termoregolazione per la vela di Dimetrodon: uno come un mezzo veloce di riscaldamento al mattino, e un altro come un mezzo di raffreddamento quando la temperatura corporea diventava troppo alta. Originariamente, Dimetrodon e tutti gli altri vertebrati terrestri del Permiano, venivano indicati come animali a sangue freddo o pecilotermi, dipendendo dal calore del sole per mantenere costante la temperatura corporea elevata, come i rettili attuali. A causa delle sue grandi dimensioni, Dimetrodon aveva un'elevata inerzia termica, il che significa che i cambiamenti nella temperatura corporea si verificavano più lentamente rispetto ad animali più piccoli. All'innalzamento della temperatura al mattino, le prede di Dimetrodon dai corpi più piccoli, si scaldavano molto più rapidamente di quanto potesse fare invece il più grande Dimetrodon. Molti paleontologi tra cui Haack hanno proposto che la vela di Dimetrodon potrebbe aver permesso di scaldarsi più rapidamente al mattino, al fine di tenere il passo con le sue prede.[44] La grande superficie della vela implicava anche che il calore si dissipava più rapidamente nell'ambiente circostante, utile se l'animale aveva bisogno di rilasciare il calore in eccesso prodotto dal metabolismo o assorbito dal sole. Dimetrodon potrebbe anche aver angolato la vela in base alla posizione del sole per rinfrescarsi o per limitare il flusso di sangue alla vela e mantenere il calore durante la notte.[42]
Nel 1986, J. Scott Turner e Richard C. Tracy hanno proposto che l'evoluzione della vela in Dimetrodon era legata all'evoluzione del sangue caldo negli antenati dei mammiferi. Turner e Tracy hanno pensato che la vela di Dimetrodon avrebbe permesso all'animale di essere omeotermo, mantenendo un costante, seppur bassa, la temperatura corporea. Anche i mammiferi sono omeotermi, anche se differiscono da Dimetrodon nell'essere endotermici, controllando la loro temperatura corporea internamente attraverso il metabolismo. Turner e Tracy notarono che i primi terapsidi, un gruppo più avanzato di sinapsidi strettamente legato ai mammiferi, hanno lunghi arti che possono rilasciare calore in un modo simile a quello della vela di Dimetrodon. L'omeotermia sviluppatosi in animali come Dimetrodon può essere riportato ai terapsidi attraverso una modifica della forma del corpo, che si sarebbe poi sviluppato nel sangue caldo dei mammiferi.[46]

Recenti studi sulla vela di Dimetrodon e di altri sfenacodontidi, supportano la tesi di Haack (1986) che affermava che la vela era scarsamente adatta a rilasciare calore e mantenere una temperatura corporea stabile. La presenza della vela nelle specie più piccole di Dimetrodon, come D. milleri e D. teutonis non si adattano all'idea che lo scopo della vela era di termoregolazione perché le vele più piccole sono insufficienti per trasferire il calore e perché i piccoli corpi possono assorbire e rilasciare calore facilmente da soli. Inoltre, i parenti più stretti di Dimetrodon, come Sphenacodon, hanno creste molto basse che sarebbero state inutili come dispositivi per la termoregolazione.[22] Si pensa che la grande vela di Dimetrodon si sia evoluta proprio da creste piccole e basse come quelle di Sphenacodon, il che significa che su gran parte della storia evolutiva della vela, la termoregolazione non sarebbe servita a molto.[47]
Gli esemplari più grandi di Dimetrodon hanno vele più grandi rispetto alle loro dimensioni, un esempio di allometria positiva. L'allometria positiva può beneficiare della termoregolazione perché significa che, come gli individui diventavano più grandi, più la superficie aumenta. Gli animali più grandi generano una grande quantità di calore attraverso il metabolismo e la quantità di calore che deve essere dissipato dalla superficie del corpo è significativamente maggiore di quello che doveva essere dissipata da animali più piccoli. La dissipazione del calore poteva essere più efficace da un singolo rapporto tra massa e superficie. Tuttavia, uno studio del 2010 di allometria in Dimetrodon ha provato un diverso rapporto tra la vela e la massa corporea: l'esponente di scala reale della vela era molto più grande dell'esponente previsto in un animale atto alla dissipazione del calore. I ricercatori hanno concluso che la vela di Dimetrodon cresceva ad un ritmo molto più veloce di quanto era necessario per la termoregolazione, e ha suggerito che la selezione sessuale è stata la ragione principale per la sua evoluzione.[47]
Selezione sessuale
[modifica | modifica wikitesto]L'esponente allometrica per l'altezza della vela è simile in grandezza alla scala di lunghezza interspecifica nelle corna all'altezza delle spalle nei cervidi. Inoltre, come Bakker (1970) ha osservata nel contesto di Dimetrodon, molte specie di lucertola sollevano una cresta di pelle dorsale durante il display di minaccia e corteggiamento, e positivamente allometriche, soprattutto nelle specie che presentano un forte dimorfismo sessuale (Echelle et al., 1978; Christian et al., 1995). Anche in Dimetrodon vi è un forte dimorfismo sessuale, soprattutto nella robustezza dello scheletro e l'altezza relativa delle spine di D. limbatus (Romer & Price, 1940).[47]
Dimorfismo sessuale
[modifica | modifica wikitesto]Anche Dimetrodon potrebbe aver avuto un forte dimorfismo sessuale, il che significa che i maschi e le femmine hanno dimensioni leggermente diverse. Alcuni esemplari di Dimetrodon sono stati ipotizzati come maschi, perché hanno ossa più spesse, vele più grandi, teschi più lunghi e mascellari più profondi. Sulla base di queste differenze, gli scheletri montati al Museo Americano di Storia Naturale (AMNH 4636) e al Field Museum sarebbero in prevalenza maschi, mentre gli scheletri del Denver Museum of Natural History (MCZ 1347) e dell'University of Michigan Museum of Natural History sarebbero femmine.[38]
-

-
 Scheletro di D. incisivus all'University of Michigan Museum of Natural History, che potrebbe rappresentare una femmina.
Scheletro di D. incisivus all'University of Michigan Museum of Natural History, che potrebbe rappresentare una femmina.


Paleoecologia
[modifica | modifica wikitesto]
I fossili di Dimetrodon sono stati ritrovati negli Stati Uniti (Texas, Oklahoma, Nuovo Messico, Arizona, Utah e Ohio) e in Germania, aree che all'epoca facevano parte del supercontinente Euramerica, durante il Permiano inferiore. Negli Stati Uniti, quasi tutto il materiale attribuito a Dimetrodon viene da tre gruppi geologici nel centro-nord del Texas e dell'Oklahoma centro-meridionale: il Gruppo Clear Fork, il Gruppo di Wichita e il Gruppo River Pearce.[48] La maggior parte dei reperti fossili fanno parte degli ecosistemi di pianura, che, durante il Permiano, sarebbero state vaste zone umide. In particolare, i Red Bed del Texas, una zona di grande diversità di fossili di tetrapodi. Oltre a Dimetrodon, i tetrapodi più comuni nel Red Bed e in tutti i depositi del Permiano presto nel sud-ovest degli Stati Uniti, sono gli anfibi Archeria, Diplocaulus, Eryops e Trimerorhachis, il rettiliomorfo Seymouria, il rettile Captorhinus e i sinapsidi Ophiacodon ed Edaphosaurus. Questi tetrapodi costituiscono un gruppo di animali che il paleontologo Everett C. Olson ha chiamato la "cronofauna del Permo-Carbonifero", una fauna che ha dominato l'ecosistema dell'Euramerica per diversi milioni di anni.[49] Sulla base della geologia dei depositi come i Red Beds, questa fauna abitava in una pianura rigogliosa e piena di vegetazione in un ecosistema deltizio.[50]
Olson ha fatto molte congetture sulla paleoecologia dei Red Beds del Texas e il ruolo di Dimetrodon all'interno del suo ecosistema. Olson ha proposto diversi tipi di ecosistema in cui i primi tetrapodi avrebbero potuto vivere. Dimetrodon apparteneva all'ecosistema più primitivo, che si è sviluppato da una catena alimentare acquatica. In esso, le piante acquatiche sono state i produttori primari e fornivano l'alimentazione principale per pesci e invertebrati acquatici. La maggior parte dei vertebrati terrestri si nutrivano principalmente di prede acquatiche. Dimetrodon era probabilmente il predatore alfa dell'ecosistema, nutrendosi di una grande varietà di organismi, come il grande squalo Xenacanthus, gli anfibi acquatici Trimerorhachis e Diplocaulus, e i tetrapodi terrestri Seymouria e Trematops. All'epoca erano presenti anche numerose specie di insetti che con buona probabilità entravano anch'essi, in una certa misura, nella catena alimentare di Dimetrodon, alimentando piccoli rettili come Captorhinus. L'assemblaggio dei Red Beds includono anche alcuni dei primi grandi erbivori terrestri come Edaphosaurus e Diadectes, che si nutrivano principalmente di prede terrestri, anziché acquatiche. Secondo Olson, il miglior analogico moderno per l'ecosistema di Dimetrodon, sono le Everglades.[50] Lo stile di vita esatto di Dimetrodon (anfibio o terrestre) è stato a lungo dibattuto, ma la microanatomia delle ossa supporta uno stile di vita terrestre,[51] il che implica che avrebbe cacciato principalmente sulla terraferma, sulle rive o in acque molto basse. Esistono anche prove di predazione di Dimetrodon ai danni di Diplocaulus, durante i periodi di siccità, grazie al ritrovamento di tre giovani Diplocaulus parzialmente mangiati in una tana e al ritrovamento di otto segni di denti recanti da un Dimetrodon, che li estrasse dal terreno e li mangiò.[52]
L'unica specie di Dimetrodon ritrovata al di fuori del sud-ovest degli Stati Uniti è D. teutonis, dalla Germania. I suoi resti sono stati ritrovati nella Formazione Tambach in un sito chiamato Località Bromacker. L'assemblaggio dei tetrapodi del Bromacker, nel Permiano inferiore è alquanto insolito, in quanto ci sono pochi grandi sinapsidi che servono il ruolo di grandi predatori. Le dimensioni di D. teutonis è stimata a soli 1,7 metri (5.6 ft) di lunghezza, troppo piccole per predare grandi diadectidi erbivori, abbondanti nell'assemblaggio del Bromacker. È più probabile che questa specie si nutrisse di piccoli vertebrati e insetti. Sono stati ritrovati solo tre fossili che potrebbero essere attribuiti a grandi predatori, e si ritiene che siano per lo più grandi o piccoli sfenacodonti varanopidi, entrambi i quali potevano potenzialmente predare D. teutonis. In contrasto con le pianura deltizie dei Red Beds del Texas, i depositi del Bromacker si pensa rappresentassero un ambiente montano con nessuna specie acquatica. È possibile che le grandi specie di Dimetrodon non fossero parte dell'ecosistema di Bromacker, perché erano dipendenti da grandi prede acquatiche.[5]
Nella cultura di massa
[modifica | modifica wikitesto]Nonostante non sia un dinosauro anche se viene più volte erroneamente identificato come tale, come animale preistorico il dimetrodonte è sorprendentemente ben conosciuto presso il grande pubblico. Spesso, poi, modellini di dimetrodonte vengono inclusi nelle confezioni di giocattoli per bambini. Nonostante la sua fama, il dimetrodonte è spesso confuso con il dinosauro Spinosaurus, a causa del fatto che entrambi questi animali presentavano una vela dorsale.
- È presente in un famoso film del 1959 con James Mason, Viaggio al centro della Terra, tratto da un romanzo di Verne, e poi è anche presente nel documentario della BBC L'impero dei mostri.
- Nel Parco della Preistoria di Rivolta d'Adda sono presenti 3 ricostruzioni di dimetrodonti. Altre ricostruzioni del dimetrodonte sono presenti al Parco Preistorico di Peccioli (PI), al Mondo della Preistoria di Simbario (VV) e al Parco dei Dinosauri di Castellana Grotte (BA).
- Era apparso anche nel film di Walt Disney Fantasia (1940) nel capitolo La sagra della primavera e compare molto brevemente anche nel primo film della serie Alla ricerca della Valle Incantata.
- Ranu, uno dei protagonisti del cartone animato I Saurini e i viaggi del meteorite nero è un dimetrodonte.
- Uno degli zord dei Power Rangers della serie Power Rangers Dino Thunder è un dimetrodonte celeste la cui "vela" è in realtà la lama di una sega circolare.
- Un giocattolo con le fattezze del dimetrodonte appare brevemente nel film d'animazione Toy Story 3 - La grande fuga prodotto dalla Pixar Animation Studios e nel corto Toy Story: Tutto un altro mondo.
- Nel franchise di Jurassic Park, il dimetrodonte compare inizialmente sotto forma di diorama all'interno della biblioteca della magione Lockwood, nel film Jurassic World - Il regno distrutto (2018). Compare fisicamente nel film Jurassic World - Il dominio (2022), dove un gruppo di questi animali attacca Alan Grant, Ellie Sattler e Maisie Lockwood in una miniera di ambra abbandonata. Insieme al Listrosauro, è uno dei primi sinapsidi a comparire all’interno del franchise.
- È apparso in videogiochi di Jurassic world come Jurassic World: il gioco, Jurassic World Alive e Jurassic World Evolution 2 nel DLC Jurassic World Dominion Biosyn Expansion con un aspetto più simile a quello del film.
Note
[modifica | modifica wikitesto]- ^ Dimetrodon, su Paleobiology Database. URL consultato il 23 agosto 2012 (archiviato dall'url originale il 2 ottobre 2013).
- ^ a b c d K. D. Angielczyk, Dimetrodon is Not a Dinosaur: Using Tree Thinking to Understand the Ancient Relatives of Mammals and their Evolution, in Evolution: Education and Outreach, vol. 2, n. 2, 2009, pp. 257-271, DOI:10.1007/s12052-009-0117-4.
- ^ Huttenlocker, A. K., and E. Rega. 2012. The Paleobiology and Bone Microstructure of Pelycosauriangrade Synapsids. Pp. 90–119 in A. Chinsamy (ed.) Forerunners of Mammals: Radiation, Histology, Biology. Indiana University Press.
- ^ Ancestry of mammalian 'warm-bloodedness' revealed, su sciencedaily.com, Society of Vertebrate Paleontology, 29 ottobre 2015. URL consultato il 29 ottobre 2015.
- ^ a b c D.S. Berman, Reisz, R.R., Martens, T. e Henrici, A.C., A new species of Dimetrodon (Synapsida: Sphenacodontidae) from the Lower Permian of Germany records first occurrence of genus outside of North America (PDF), in Canadian Journal of Earth Sciences, vol. 38, n. 5, 2001, pp. 803-812, DOI:10.1139/cjes-38-5-803.
- ^ The Kungurian Age, su Palaeos. URL consultato il 29 settembre 2013 (archiviato dall'url originale il 12 novembre 2013).
- ^ a b c J. Fröbisch, Schoch, R.R., Müller, J., Schindler, T. e Schweiss, D., A new basal sphenacodontid synapsid from the Late Carboniferous of the Saar-Nahe Basin, Germany (PDF), in Acta Palaeontologica Polonica, vol. 56, n. 1, 2011, pp. 113-120, DOI:10.4202/app.2010.0039.
- ^ E.C. Olson e Beerbower, J.R., The San Angelo Formation, Permian of Texas, and its vertebrates, in The Journal of Geology, vol. 61, n. 5, 1953, pp. 389-423, Bibcode:1953JG.....61..389O, DOI:10.1086/626109.
- ^ E.C. Olson, Parallelism in the evolution of the Permian reptilian faunas of the Old and New Worlds, in Fieldiana, vol. 37, n. 13, 1955, pp. 385-401.
- ^ C.W. Sternberg, The skeleton of an immature pelycosaur, Dimetrodon cf. grandis, from the Permian of Texas, in Journal of Paleontology, vol. 16, n. 4, 1942, pp. 485-486, JSTOR 1298848.
- ^ M. Laurin e Reisz, R.R., Synapsida: mammals and their extinct relatives, su Tree of Life Web Project, 2012. URL consultato il 24 agosto 2012 (archiviato dall'url originale il 20 gennaio 2013).
- ^ Exhibit Specimens: Dimetrodon, su American Museum of Natural History. URL consultato il 2 luglio 2012 (archiviato dall'url originale il 4 luglio 2012).
- ^ a b c G. Baur e Case, E.C., The history of the Pelycosauria, with a description of the genus Dimetrodon, Cope, in Transactions of the American Philosophical Society, vol. 20, n. 1, 1899, pp. 5-62, JSTOR 1005488.
- ^ a b c Abler, W.L. 2001. A kerf-and-drill model of tyrannosaur tooth serrations. p. 84-89. In: Mesozoic Vertebrate Life. Ed.s Tanke, D. H., Carpenter, K., Skrepnick, M. W. Indiana University Press.
- ^ http://phenomena.nationalgeographic.com/2014/02/07/sail-backed-dimetrodon-had-a-nasty-bite/
- ^ T.S. Kemp, The origin and early radiation of the therapsid mammal-like reptiles: a palaeobiological hypothesis, in Journal of Evolutionary Biology, vol. 19, n. 4, 2006, pp. 1231-1247, DOI:10.1111/j.1420-9101.2005.01076.x, PMID 16780524.
- ^ M. Laurin e Reisz, R.R., Autapomorphies of the main clades of synapsids, su Tree of Life Web Project, 1997. URL consultato il 24 agosto 2012 (archiviato dall'url originale l'11 dicembre 2008).
- ^ A.S. Romer, Notes on the Permo-Carboniferous reptile Dimetrodon, in The Journal of Geology, vol. 35, n. 8, 1927, pp. 673-689, Bibcode:1927JG.....35..673R, DOI:10.1086/623462, JSTOR 30060393.
- ^ a b E. A. Rega, K. Noriega, S. S. Sumida, A. Huttenlocker, A. Lee e B. Kennedy, Healed Fractures in the Neural Spines of an Associated Skeleton of Dimetrodon: Implications for Dorsal Sail Morphology and Function, in Fieldiana Life and Earth Sciences, vol. 5, 2012, pp. 104-111, DOI:10.3158/2158-5520-5.1.104.
- ^ a b c E. Rega, Sumida, S., Noriega, K., Pell, C. e Lee, A., Evidence-based paleopathology I: Ontogenetic and functional implications of dorsal sails in Dimetrodon, in Journal of Vertebrate Paleontology, vol. 25, S3, 2005, pp. 103A, DOI:10.1080/02724634.2005.10009942.
- ^ S. Sumida, Rega, E. e Noriega, K., Evidence-based paleopathology II: Impact on phylogenetic analysis of the genus Dimetrodon, in Journal of Vertebrate Paleontology, vol. 25, S3, 2005, pp. 120A, DOI:10.1080/02724634.2005.10009942.
- ^ a b c A.K. Huttenlocker, Rega, E. e Sumida, S.S., Comparative anatomy and osteohistology of hyperelongate neural spines in the sphenacodontids Sphenacodon and Dimetrodon (Amniota: Synapsida), in Journal of Morphology, vol. 271, n. 12, 2010, pp. 1407-1421, DOI:10.1002/jmor.10876, PMID 20886514.
- ^ E.A. Rega, Noriega, K., Sumida, S. e Lee, A., Histological analysis of traumatic injury to multiple neural spines of an associated skeleton of Dimetrodon: Implications for healing response, dorsal sail morphology and age-at-death in a Lower Permian synapsid, in Integrated Comparative Biology, vol. 44, 2004, p. 628.
- ^ Petr Konstantinovich Tchudinov, New Facts about the Fauna of the Upper Permian of the USSR, in Journal of Geology, vol. 73, 1965, pp. 117-30.
- ^ Botha-Brink, J.; Modesto, S.P. (2007). "A mixed-age classed 'pelycosaur' aggregation from South Africa: earliest evidence of parental care in amniotes?". Proceedings of the Royal Society B 274 (1627): 2829–2834. doi:10.1098/rspb.2007.0803. PMC 2288685. PMID 17848370.
- ^ Niedźwiedzki, G.; Bojanowski, M. (2012). "A Supposed Eupelycosaur Body Impression from the Early Permian of the Intra-Sudetic Basin, Poland". Ichnos 19 (3): 150–155. doi:10.1080/10420940.2012.702549
- ^ W.D. Matthew, Review of Case's 'Revision of the Pelycosauria of North America', in Science, vol. 27, n. 699, 1908, pp. 816-818, DOI:10.1126/science.27.699.816.
- ^ a b E.C. Case, Revision of the Pelycosauria of North America, Washington, D.C., Carnegie Institution of Washington, 1907, pp. 1-176.
- ^ E.D. Cope, Descriptions of extinct Batrachia and Reptilia from the Permian formation of Texas, in Proceedings of the American Philosophical Society, vol. 17, n. 101, 1878, pp. 505–530, JSTOR 982652.
- ^ E.D. Cope, Descriptions of extinct vertebrata from the Permian and Triassic Formations of the United States, in Proceedings of the American Philosophical Society, vol. 17, n. 100, 1877, pp. 182–193, JSTOR 982295.
- ^ P.P. Vaughn, Comparison of the Early Permian vertebrate faunas of the Four Corners region and north-central Texas (PDF), Contributions in Science, vol. 105, Los Angeles County Museum of Natural History, 1966, pp. 1-13. URL consultato il 22 marzo 2017 (archiviato dall'url originale il 10 gennaio 2017).
- ^ P.P. Vaughn, Early Permian vertebrates from southern New Mexico and their paleozoogeographic significance (PDF), Contributions in Science, vol. 166, Los Angeles County Museum of Natural History, 1969, pp. 1-22. URL consultato il 22 marzo 2017 (archiviato dall'url originale il 10 gennaio 2017).
- ^ E.C. Olson, Permo-Carboniferous paleoecology and morphotypic series, in American Zoologist, vol. 15, n. 2, 1975, pp. 371-389, DOI:10.1093/icb/15.2.371.
- ^ a b D.S. Berman, A new species of Dimetrodon (Reptilia, Pelycosauria) from a non-deltaic facies in the Lower Permian of north-central New Mexico, in Journal of Paleontology, vol. 51, n. 1, 1977, pp. 108-115, JSTOR 1303466.
- ^ K. Madalena, Sumida, S., Zeigler, K. e Rega, E., A new record of the Early Permian pelycosaurian-grade synapsid Dimetrodon (Eupelycosauria: Sphenacodontidae) from the Lower Cutler Group (Early Permian) of Jemez Pueblo, north-central New Mexico, in Journal of Vertebrate Paleontology, vol. 27, 3, Suppl., 2007, pp. 110A, DOI:10.1080/02724634.2007.10010458.
- ^ P.P. Vaughn, Vertebrates from the Cutler Group of Monument Valley and vicinity (PDF), New Mexico Geological Society Guidebook, vol. 24, New Mexico Geological Society, 1973, pp. 99-105. URL consultato il 22 marzo 2017 (archiviato dall'url originale il 12 aprile 2016).
- ^ The mammals appear
- ^ a b A.S. Romer e Price, L.I., Review of the Pelycosauria, in Geological Society of America Special Paper, Geological Society of America Special Papers, vol. 28, 1940, pp. 1-538, DOI:10.1130/spe28-p1.
- ^ a b R.J. Benson, Interrelationships of basal synapsids: cranial and postcranial morphological partitions suggest different topologies, in Journal of Systematic Paleontology, vol. 10, n. 2, 2012, pp. 601-624, DOI:10.1080/14772019.2011.631042.
- ^ R. R. Reisz, Pelycosauria, in Sues, H.-D. (ed.) (a cura di), Handbuch der Paläoherpetologie, 17A, Gustav Fischer Verlag, 1986, pp. 1-102, ISBN 978-3-89937-032-4, OL 12985656M.
- ^ Kirstin S. Brink, Hillary C. Maddin, David C. Evans, Robert R. Reisz e Hans-Dieter Sues, Re-evaluation of the historic Canadian fossil Bathygnathus borealisfrom the Early Permian of Prince Edward Island, in Canadian Journal of Earth Sciences, vol. 52, n. 12, 2015, pp. 1109-1120, DOI:10.1139/cjes-2015-0100.
- ^ a b G.A. Florides, Kalogirou, S.A., Tassou, S.A. e Wrobel, L., Natural environment and thermal behaviour of Dimetrodon limbatus, in Journal of Thermal Biology, vol. 26, n. 1, 2001, pp. 15-20, DOI:10.1016/S0306-4565(00)00019-X, PMID 11070340.
- ^ C.D. Bramwell e Fellgett, P.B., Thermal regulation in sail lizards (PDF), in Nature, vol. 242, n. 5394, 1973, pp. 203-205, DOI:10.1038/242203a0.
- ^ a b S.C. Haack, A thermal model of the sailback pelycosaur, in Paleobiology, vol. 12, n. 4, 1986, pp. 450-458.
- ^ G.A. Florides, Wrobel, L.C., Kalogirou, S.A. e Tassou, S.A., A thermal model for reptiles and pelycosaurs, in Journal of Thermal Biology, vol. 24, n. 1, 1999, pp. 1-13, DOI:10.1016/S0306-4565(98)00032-1.
- ^ J.S. Turner e Tracy, C.R., Body size, homeothermy and the control of heat exchange in mammal-like reptiles (PDF), in Hotton, N. III.; MacLean, P.D.; Roth, J.J.; and Roth, E.C. (eds.) (a cura di), The Ecology and Biology of Mammal-Like Reptiles, Washington, D.C., Smithsonian Institution Press, 1986, pp. 185-194. URL consultato il 23 marzo 2017 (archiviato dall'url originale il 12 aprile 2016).
- ^ a b c J.L. Tomkins, LeBas, N.R., Witton, M.P., Martill, D.M. e Humphries, S., Positive allometry and the prehistory of sexual selection (PDF), in The American Naturalist, vol. 176, n. 2, 2010, pp. 141-148, DOI:10.1086/653001, PMID 20565262 (archiviato dall'url originale il 12 aprile 2016).
- ^ S.G. Lucas, Spielmann, J.A., Rinehart, L.F. e Martens, T., Dimetrodon (Amniota: Synapsida: Sphenacodontidae) from the Lower Permian Abo Formation, Socorro County, New Mexico (PDF), New Mexico Geological Society Guidebook, vol. 60, New Mexico Geological Society, 2009, pp. 281-284. URL consultato il 23 marzo 2017 (archiviato dall'url originale il 5 ottobre 2021).
- ^ C. Sullivan, Reisz, R.R. e May, W.J., [0456:LDSEFT2.0.CO;2 Large dissorophoid skeletal elements from the Lower Permian Richards Spur fissures, Oklahoma, and their paleoecological implications], in Journal of Vertebrate Paleontology, vol. 20, n. 3, 2000, pp. 456-461, DOI:10.1671/0272-4634(2000)020[0456:LDSEFT]2.0.CO;2, JSTOR 4524117.
- ^ a b E.C. Olson, Community evolution and the origin of mammals, in Ecology, vol. 47, n. 2, 1966, pp. 291-302, DOI:10.2307/1933776, JSTOR 1933776.
- ^ A. Kriloff, Germain, D., Canoville, A., Vincent, P., Sache, M. e Laurin, M., Evolution of bone microanatomy of the tetrapod tibia and its use in palaeobiological inference, in Journal of Evolutionary Biology, vol. 21, n. 3, 2008, pp. 807-826, DOI:10.1111/j.1420-9101.2008.01512.x, PMID 18312321.
- ^ http://www.livescience.com/40842-finned-monster-chomped-heads-off-ancient-amphibians.html
Bibliografia
[modifica | modifica wikitesto]- Bramwell, C. D. and Fellgett, P. P., 1973, Termoregolazione nelle "lucertole" munite di vela. Nature, v. 242, p. 203-205
Altri progetti
[modifica | modifica wikitesto] Wikimedia Commons contiene immagini o altri file su Dimetrodon
Wikimedia Commons contiene immagini o altri file su Dimetrodon Wikispecies contiene informazioni su Dimetrodon
Wikispecies contiene informazioni su Dimetrodon
Collegamenti esterni
[modifica | modifica wikitesto]- Dimetrodon, su sapere.it, De Agostini.
- (EN) Dimetrodon, su Enciclopedia Britannica, Encyclopædia Britannica, Inc.
- (EN) Dimetrodon, su Fossilworks.org.